The Power of Unity: Understanding Aggregation Pheromones
Abstract: Aggregation pheromones are a class of chemical signals that promote the mass clustering of conspecifics at a particular location. Unlike sex pheromones, which are…
Read moreDive into daily dispatches exploring neuroscience, emerging tech, climate breakthroughs, and the stories shaping tomorrow.
Abstract: Aggregation pheromones are a class of chemical signals that promote the mass clustering of conspecifics at a particular location. Unlike sex pheromones, which are…
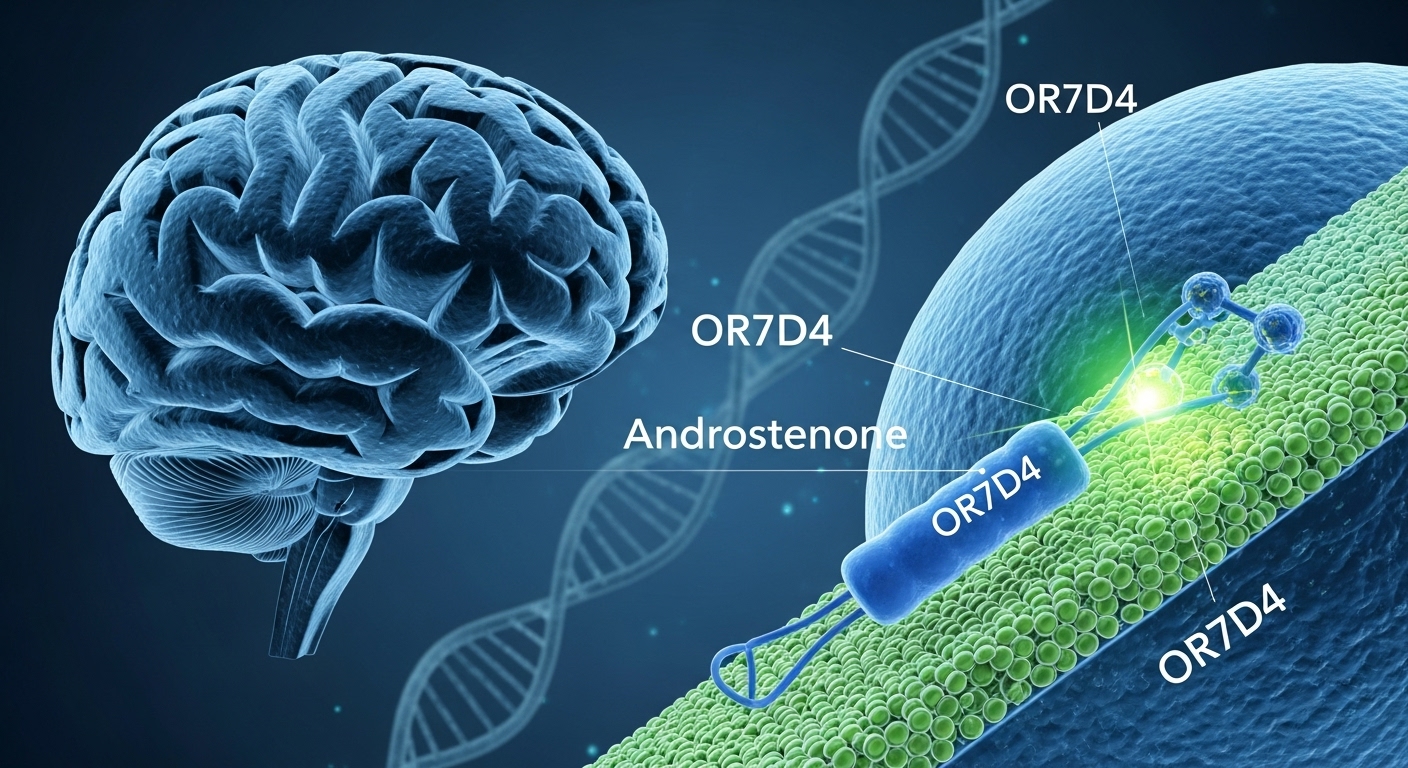
Read moreAbstract: The existence of human pheromones remains a topic of scientific controversy, but recent genetic research has provided the clearest link yet between specific odorant…
Read moreAbstract: Pheromones are broadly categorized by the nature and duration of the response they elicit in the receiver. The two primary functional classes are *releaser*…
Read moreAbstract: Alarm pheromones are a critical class of semiochemicals that induce immediate, stereotypical defensive or evasive behaviors in conspecifics upon exposure to threat. Unlike sex…
Read moreAbstract: Sex pheromones, particularly in moths (Lepidoptera), are arguably the best-understood chemical communication systems in nature, serving as an essential long-distance signaling mechanism for reproduction.…
Read moreAbstract: Trail pheromones represent one of the most efficient forms of chemical communication in the animal kingdom, particularly among social insects like ants. These volatile…
Read moreIt's a widely held belief, shared among friends and passed down through generations: when women live in close proximity, their menstrual cycles align. This phenomenon,…
Read moreWhen you think of pheromones, you might imagine a moth searching for a mate in the dark. But what if that same powerful attraction could…
Read moreThe desire to capture affection and inspire passion is as old as humanity itself. Long before the advent of modern chemistry, ancient Greeks and Romans…
Read more